Cell polarity and cell fate in the Drosophila female and male reproductive system: a Cell Biological and Biophysical approach
Research description
Our research group focuses on understanding the genetic, cell biology and biophysics basis of cell polarity and cell fate in a multicellular organism. Experimental biophysicists and biologists are welcome to join us.
We use Drosophila melanogaster as a model system to understand how cells adopt their fate in both the female and male germline, and how the oocyte is polarised. These are basic cellular processes that are conserved in the development of all organisms. More specifically, the lab has two main lines of research:
1. Cytoplasmic flows and cellular asymmetries in the oocyte
We are investigating cellular asymmetries in the oocyte, and their relation with the fluid mechanical properties of the cytoplasm. In collaboration with physicist Prof. R. Goldstein, we have a unique approach that combines interdisciplinary experimental and theoretical parameters in order to answer these questions. A physicist (Dr. A. Jimenez Dalmaroni) is soon joining the lab. In the oocyte, as the developmental determinants are being asymmetrically localised, motor proteins also induce the vigorous movement of the oocyte cytoplasm, known as cytoplasmic streaming. Cytoplasmic streaming was discovered in 1774, but many fundamental questions have remained unanswered: How does the fluid motion arise? What is the relationship between the oocyte asymmetries and the underlying forces of the observed flows? We have engaged in a comprehensive experimental and theoretical study of fluid dynamical and transport issues, using techniques from microfluidics to functional genetics. This interdisciplinary approach allows us to cover the various aspects of quantitative biological data collection, physical modelling and experimental design required to study the relation between the fluid mechanical properties of the cytoplasm, the cytoskeletons (actin and microtubules) and oocyte polarity. We are also studying how motors achieve specificity in their functions. This work combines traditional genetic and cell biological approaches with state-of-the art analysis of single molecule dynamics.
2. Regulation of cell fate and tissue architecture in ovaries and testis
The second line of research examines the function of the conserved Hippo (Hpo) tumour-suppressor pathway in cell fate and tissue architecture. In brief, the Hpo pathway consists of a cascade of kinases that regulate the transcription of cell growth genes. We (and others) have showed that Hpo also influences cell fate and tissue architecture. However, not much is known about these Hpo functions. Although many upstream regulators of Hpo are characterised, the identity and function of its transcriptional targets are largely unknown. We are addressing these questions combining functional genomic and cell biology approaches.
2.1. Hippo and cell differentiation in the female germline. The polarisation of the oocyte (required for the localisation of the developmental determinants) relies on the Hpo-dependent differentiation of a group of cells, the posterior follicle cells (FCs), which are in direct contact with the oocyte. We aim to understand what makes the posterior FCs become posterior, and how Hpo contributes to this process. To achieve this, we have obtained the genomic profiling of posterior FCs, and we have compared it to the transcriptome in other FCs. We are currently analysing candidate genes, and the preliminary results are very encouraging. In addition, in contrast to wildtype cells, hpo mutant FCs form a multilayered epithelium. We are investigating how the follicular epithelium maintains a monolayer, and how this mechanism is disrupted in hpo mutant cells.
2.2. Hippo and cell differentiation in the male germline. We have recently found that Hpo also controls the fate of the germline stem cells in testis (GSCs). Stem cells are crucial for homeostasis (maintaining populations of highly differentiated cell types), and thus a balance between self-renewal and differentiation must be maintained. Signals from the niche cells (of somatic origin) regulate this balance. However, little is known about how niche cell number and function is regulated. We have observed that GSCs can lose their stem cell-ness and incorporate into the niche as a niche cell. This is rarely observed in young testis, but it is more prominent in older testis. These observations raise the exciting possibility that GSCs contribute to maintaining a “healthy” niche during the known age-related decline in niche function. We have also found that mutating the Hpo pathway in GSCs dramatically increases the events of GSC-to-niche transformation. We aim to further study this novel switch in stem cell fate, and how Hpo regulates it.
Our movies
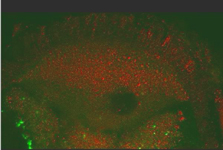
Movie 1. Movement of GFP-Staufen and red particles in a wild type egg chamber
The movement of GFP-Staufen (an excellent marker for oskar mRNA) and red particles (that reflect the 568nm light) in a wild type egg chamber were analyzed by time lapse films on a BioRad Confocal MRC1024. The time lapse movie was obtained by collecting Z series of 5 sections at 2 μm intervals every minute for 45 minutes. This movie lasts 8 seconds and represents 4 consecutive loops of one single movie (thus, 45 mins of real time are shown in a 2 seconds movie). The posterior pole of the oocyte is at the bottom of the image.
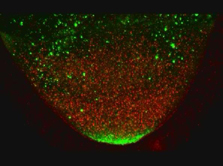
Movie 2. Movement of GFP-Staufen and red particles in a kinesin heavy chain mutant egg chamber
Kinesin Heavy Chain is essential for the induction of cytoplasmic flows within the oocyte. The movement of GFP-Staufen (an excellent marker for oskar mRNA) and red particles (that reflect the 568nm light) in a kinesin heavy chain egg chamber were analyzed by time lapse films on a BioRad Confocal MRC1024. The time lapse movie was obtained by collecting Z series of 5 sections at 2 μm intervals every minute for 45 minutes. This movie lasts 8 seconds and represents 4 consecutive loops of one single movie (thus, 45 mins of real time are shown in a 2 seconds movie). The posterior pole of the oocyte is at the top of the image